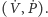O pewnym ciekawym zastosowaniu modelu drapieżnik–ofiara
W artykule tym podejmiemy próbę wyjaśnienia nietypowej dysproporcji gatunkowej na kontynencie australijskim. Chodzi o niespotykany nigdzie indziej brak stałocieplnych drapieżników, przy jednoczesnym rozkwicie zimnokrwistych mięsożerców. Zwrócił na to uwagę w swoim artykule The case of missing meat eaters (opublikowanym w Natural History w 1993 roku) Tim Flannery, mammolog i paleontolog, specjalizujący się w australijskim ekosystemie.

Powodów zaistniałego stanu rzeczy doszukiwał się w plejstocenie, gdy człowiek
jeszcze nie dotarł na kontynent, a Australia wraz z przyległymi wyspami
tworzyła tzw. Meganezję. W tym czasie na wszystkich kontynentach żyło wiele
wymarłych dziś gatunków, należących do tzw. megafauny, czyli populacji
zwierząt, których masa często przekraczała tonę, jak np. mamut włochaty,
który osiągał masę
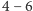 ton, czy Indrikoterium, nieparzystokopytny ssak
roślinożerny, blisko spokrewniony z nosorożcowatymi, o masie do 15 ton.
Pod koniec plejstocenu wielkie wymieranie dotknęło zwierzęta z całego globu,
ale wszędzie – poza Australią – przetrwało wiele gatunków ssaków, w tym
drapieżnych, osiągających dużą masę, w szczególności różne gatunki
kotowatych (puma, ryś, tygrys, lampart, lew, jaguar i gepard).
ton, czy Indrikoterium, nieparzystokopytny ssak
roślinożerny, blisko spokrewniony z nosorożcowatymi, o masie do 15 ton.
Pod koniec plejstocenu wielkie wymieranie dotknęło zwierzęta z całego globu,
ale wszędzie – poza Australią – przetrwało wiele gatunków ssaków, w tym
drapieżnych, osiągających dużą masę, w szczególności różne gatunki
kotowatych (puma, ryś, tygrys, lampart, lew, jaguar i gepard).
Jako jedną z możliwych przyczyn wymarcia australijskiej megafauny, wskazywaną przez wielu biologów, Flannery przytoczył niewielkie rozmiary Meganezji, co miało skutkować zbyt małą przestrzenią życiową. Jednakże np. Madagaskar wykazuje porównywalne zróżnicowanie gatunkowe, choć jego obszar jest ponad dwadzieścia razy mniejszy.
Kolejną często przytaczaną hipotezą jest stwierdzenie, że torbacze miały relatywnie małe mózgi, co uniemożliwiło im wyewoluowanie w skuteczne drapieżniki. Jednak np. z czwartorzędowych południowoamerykańskich drapieżnych torbaczy wyewoluowały drapieżniki przypominające północnoamerykańskie tygrysy szablozębne, zdolne do skutecznego polowania na największe ówcześnie żyjące zwierzęta roślinożerne. Podobnie przodkowie amerykańskiego oposa dali początek wielu przedstawicielom megafauny.
Bardziej prawdopodobna wydaje się inna przyczyna. Australia jest starym kontynentem, o stabilnej historii geologicznej. Przez ostatnie 50 milionów lat nie doświadczała zlodowaceń, procesów górotwórczych, czy aktywności wulkanicznej – procesów niezbędnych przy tworzeniu gleb. Ponadto zwarta linia brzegowa, brak większych zatok, wysoka krawędź Wyżyny Zachodnioaustralijskiej oraz Wielkie Góry Wododziałowe położone na wschodzie sprawiają, że przenikanie wpływów oceanicznych do wnętrza kontynentu jest niewielkie. Dlatego na większości terenów Australia jest wyjątkowo nieurodzajna – panuje tam suchy klimat kontynentalny, bywa, że pora deszczowa nie nadchodzi przez kilka lat z rzędu, a szata roślinna tworzy przede wszystkim stepy, półpustynie i pustynie. Dzieje się tak pomimo tego, że Australia leży na południowej półkuli, gdzie przewaga powierzchni mórz nad lądami jest znaczna, co teoretycznie powinno istotnie łagodzić i stabilizować klimat.
Ten ciągły nieurodzaj powoduje, że australijscy roślinożercy są zmuszeni
żyć w dużo większym rozproszeniu niż roślinożercy żyjący na innych
kontynentach. Jak pisze Flannery, zmniejszona liczebność potencjalnych ofiar
sprawia, że tylko populacje mięsożerców, które mają odpowiednio małe
zapotrzebowanie na pożywienie, są w stanie przetrwać. Wobec tego wśród
drapieżników faworyzowane są te o mniejszych rozmiarach ciała albo
o wolniejszym metabolizmie – w obu przypadkach do przeżycia potrzeba
mniejszych ilości pożywienia. Kręgowce zmiennocieplne mają ponad sześć
razy mniejsze zapotrzebowanie na energię niż torbacze, a dziesięć razy
mniejsze niż łożyskowce. Oznacza to, że największy znany drapieżny
torbacz, lew workowaty, potrzebował sześć razy więcej upolowanych ofiar niż
konkurujące z nim Quinkana (krokodyle ważące ponad
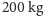 ),
Wonambi (węże ważące 50
),
Wonambi (węże ważące 50
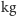 ) czy Megalania (spokrewnione
z waranem jaszczurki, dwa razy większe niż współczesne, mierzące
) czy Megalania (spokrewnione
z waranem jaszczurki, dwa razy większe niż współczesne, mierzące
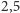 do 3 metrów warany z Komodo). Ponadto, krokodyle, węże
i jaszczurki, ponieważ nie muszą utrzymywać stałej temperatury ciała,
potrafią przetrwać bez pokarmu znacznie dłużej niż zwierzęta stałocieplne,
co przy trudnym australijskim klimacie jest dodatkową zaletą.
do 3 metrów warany z Komodo). Ponadto, krokodyle, węże
i jaszczurki, ponieważ nie muszą utrzymywać stałej temperatury ciała,
potrafią przetrwać bez pokarmu znacznie dłużej niż zwierzęta stałocieplne,
co przy trudnym australijskim klimacie jest dodatkową zaletą.
Gady, takie jak Quinkana, Wonambi czy Megalania, wyginęły w plejstocenie,
podobnie jak lew workowaty i wiele innych zwierząt megafauny. Jednak
potomkowie gadzich olbrzymów, jak waran z Komodo, nadal żyją, natomiast
większość torbaczy wówczas bezpowrotnie zniknęła. Wyginęły wszystkie
drapieżniki osiągające więcej niż 5
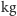 , wyłączając diabła tasmańskiego
i wilka workowatego.
, wyłączając diabła tasmańskiego
i wilka workowatego.
Właśnie to zagadnienie, czyli dynamikę populacji drapieżnika przy ograniczonej zasobności ofiar, przeanalizujemy za pomocą modelu Lotki–Volterry z ograniczoną pojemnością środowiska dla ofiar. Analiza ta ma na celu zweryfikowanie przypuszczeń Flannery’ego co do powodów zniknięcia drapieżnych torbaczy.

Model matematyczny
Zastosowany przez nas model matematyczny bazuje na klasycznym modelu
Lotki–Volterry, który jest najstarszym znanym modelem opisującym interakcje
między dwiema populacjami. Został on użyty do opisu dynamiki populacji ryb
w Adriatyku przez Volterrę w 1926 roku. Rybacy łowiący ryby w Adriatyku
zauważyli, że w czasie trwania i niedługo po zakończeniu pierwszej
wojny światowej populacja ryb drapieżnych w Morzu Śródziemnym
znacznie wzrosła. Ówcześni biolodzy nie potrafili wyjaśnić tego,
zdawało im się, paradoksalnego zjawiska. Volterra w swojej pracy Variazioni
e fluttuazioni del numero d’individui in specie animali conviventi zaproponował
model, za pomocą którego wyjaśnił, dlaczego wstrzymanie połowów
spowodowało ten nagły wzrost. Nieco wcześniej, w 1920 roku, Lotka
niezależnie od Volterry, skonstruował ten sam model i użył go do opisu
zmian stężeń dwóch reagujących ze sobą substancji chemicznych.
Obecnie najbardziej znanym przykładem zastosowania tego modelu jest
analiza zmian populacji kanadyjskich rysi i zajęcy, wykonana na podstawie
danych z lat
 dotyczących skupu skór przez Kompanię
Zatoki Hudsona.
dotyczących skupu skór przez Kompanię
Zatoki Hudsona.
Aby sformułować model, musimy przyjąć pewne założenia dotyczące
opisywanych populacji i ich środowiska. Niech
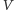 oznacza populację
ofiar, zaś
oznacza populację
ofiar, zaś
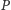 populację drapieżników (poniżej te same oznaczenia
zostaną użyte do opisania zagęszczeń populacji ofiar i drapieżników
odpowiednio – nie powinno to prowadzić do niejasności).
populację drapieżników (poniżej te same oznaczenia
zostaną użyte do opisania zagęszczeń populacji ofiar i drapieżników
odpowiednio – nie powinno to prowadzić do niejasności).
- 1.
- W środowisku występują wyłącznie rozważane
gatunki
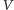 oraz
oraz
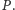 Innych zwierząt w danym środowisku
nie ma, lub nie mają wpływu na badaną dynamikę populacji, zarówno
drapieżników
Innych zwierząt w danym środowisku
nie ma, lub nie mają wpływu na badaną dynamikę populacji, zarówno
drapieżników
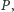 jak i ofiar
jak i ofiar
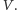
- 2.
- Gdy nie ma drapieżników,
środowisko jest sprzyjające dla ofiar – nie ma w nim żadnych innych
ograniczeń. W takiej sytuacji ich dynamika jest opisywana równaniem
Malthusa (wzrostu wykładniczego):
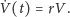
- 3.
- Gdy nie ma ofiar, drapieżniki nie mają pożywienia, co powoduje
wymieranie gatunku:
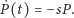
- 4.
- Ofiary rozmnażają się niezależnie od polowań drapieżników.
Drapieżniki polując na ofiary, zmniejszają ich populację, zatem wzrost
populacji
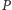 powoduje spadek
populacji
powoduje spadek
populacji
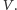 Jednocześnie drapieżniki
zdobywają pokarm, który dostarcza im energię potrzebną do życia,
w tym do rozmnażania się – zatem populacja
Jednocześnie drapieżniki
zdobywają pokarm, który dostarcza im energię potrzebną do życia,
w tym do rozmnażania się – zatem populacja
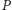 wzrasta wraz
ze wzrostem populacji
wzrasta wraz
ze wzrostem populacji
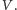
Zakładając dodatkowo równomierne rozmieszczenie osobników w przestrzeni, otrzymujemy klasyczny model Lotki–Volterry, opisany następującym układem równań różniczkowych zwyczajnych:
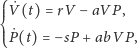 | (1) |
gdzie:
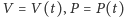 to odpowiednio zagęszczenie populacji ofiar
drapieżników (zmienne względem czasu
to odpowiednio zagęszczenie populacji ofiar
drapieżników (zmienne względem czasu
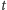 ),
),
 – współczynnik
rozrodczości ofiar,
– współczynnik
rozrodczości ofiar,
 – skuteczność polowań,
– skuteczność polowań,
 – śmiertelność
w populacji drapieżników i
– śmiertelność
w populacji drapieżników i
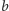 to część biomasy przeznaczona
na reprodukcję, przy czym wszystkie powyższe współczynniki są dodatnie.
to część biomasy przeznaczona
na reprodukcję, przy czym wszystkie powyższe współczynniki są dodatnie.
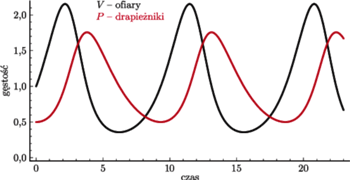
Rys. 1

Rys. 2
Analiza tego modelu pozwala uzyskać portret fazowy i przebieg rozwiązań, jak
przedstawiono na rysunkach. Rysunek 1. to wykres gęstości populacji
drapieżników i ofiar, rysunek 2. zaś to portret fazowy klasycznego modelu
Lotki–Volterry, opisanego układem (1). Wyróżniony cykl odpowiada
warunkowi początkowemu
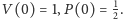
Faktycznie, jeśli w układzie (1) uwzględnimy odławianie, to dostaniemy:

gdzie
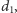
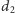 są współczynnikami odławiania odpowiednio ofiar
i drapieżników. Przy założeniu, że
są współczynnikami odławiania odpowiednio ofiar
i drapieżników. Przy założeniu, że
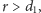 czyli odłowy nie prowadzą
do zagłady gatunku
czyli odłowy nie prowadzą
do zagłady gatunku
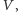 dostajemy układ Lotki–Volterry ze zmienionymi
współczynnikami:
dostajemy układ Lotki–Volterry ze zmienionymi
współczynnikami:

czyli odławianie działa zawsze na niekorzyść drapieżników, a na korzyść ofiar. Zauważmy, że z tej prostej zasady wynika też, iż warto się zastanowić ingerując w układy ekologiczne, w których ofiara jest jakimś szkodnikiem (np. populacją dokuczliwych owadów), bo poskutkuje to głównie zmniejszeniem populacji drapieżników, które z naszego punktu widzenia są pożyteczne. Oczywiście, jeśli wytępimy gatunek ofiar, to zginie także gatunek drapieżników.
Kiedy populacja ofiar się rozwija, drapieżniki mają więcej pożywienia, więc ich
liczność również wzrasta. Jednakże im więcej drapieżników, tym więcej
odławianych przez nie ofiar, a zatem po pewnym czasie liczebność populacji
ofiar zmaleje. To z kolei powoduje zmniejszenie dostępności pożywienia dla
drapieżników, a więc spadek ich liczebności, a to z konieczności
zmniejsza odsetek upolowanych ofiar i umożliwia wzrost ich liczby – tak cykl
się zamyka. Rysunki wykonane zostały przy stałych równych odpowiednio:
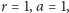
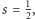
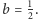
Widzimy też, że rozwiązania są okresowe, o maksimach przesuniętych
w czasie, co dobrze oddaje cykliczność zjawisk zachodzących w przyrodzie.
Co więcej, rozwiązania oscylują wokół dodatniego stanu stacjonarnego
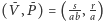 i ich wartości średnie są równe współrzędnym tego
punktu niezależnie od trajektorii, czyli także od warunku początkowego.
Własność tę nazywamy prawem zachowania średnich w układzie
drapieżnik–ofiara i jest ona przyczyną zmian zaobserwowanych przez rybaków
po pierwszej wojnie światowej.
i ich wartości średnie są równe współrzędnym tego
punktu niezależnie od trajektorii, czyli także od warunku początkowego.
Własność tę nazywamy prawem zachowania średnich w układzie
drapieżnik–ofiara i jest ona przyczyną zmian zaobserwowanych przez rybaków
po pierwszej wojnie światowej.
Jak widzimy, nawet tak prosty model, jak opisany powyżej, może przyczynić
się do wyjaśnienia pewnych zjawisk biologicznych. Jednak mimo tych zalet
model Lotki–Volterry nie jest pozbawiony wad, nie uwzględnia on bowiem
wielu zjawisk mających istotny wpływ na dynamikę populacji, jak chociażby
konkurencji o pokarm pomiędzy ofiarami, gdy ich populacja wzrasta. Dałoby
to pewne ograniczenie górne, a tym samym hamulec wzrostu wykładniczego
ofiar. Ponieważ ten właśnie czynnik ma, według Flannery’ego, zasadnicze
znaczenie w środowisku australijskim, więc przedstawimy modyfikację
układu (1) uwzględniającą pojemność środowiska dla populacji ofiar.
Do równania opisującego populację ofiar w modelu (1) dodamy składnik
postaci
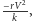 odpowiadający wewnątrzgatunkowej konkurencji
o pożywienie, gdzie współczynnik
odpowiadający wewnątrzgatunkowej konkurencji
o pożywienie, gdzie współczynnik
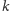 oznacza pojemność
środowiska. Składnik ten można także interpretować w następujący
sposób. Rozważmy model dynamiki populacji ofiar w przypadku
braku drapieżników:
oznacza pojemność
środowiska. Składnik ten można także interpretować w następujący
sposób. Rozważmy model dynamiki populacji ofiar w przypadku
braku drapieżników:

gdzie mamy
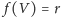 przy wykładniczym wzroście populacji, natomiast
jeśli założymy, że populacja rozwija się w ograniczonym środowisku, to
oczywiste wydaje się założenie, że
przy wykładniczym wzroście populacji, natomiast
jeśli założymy, że populacja rozwija się w ograniczonym środowisku, to
oczywiste wydaje się założenie, że
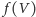 powinna być malejącą
funkcją
powinna być malejącą
funkcją
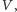 gdyż im więcej jest osobników w środowisku, tym wolniej
może się ta populacja rozwijać. Musi także być taka wielkość
populacji
gdyż im więcej jest osobników w środowisku, tym wolniej
może się ta populacja rozwijać. Musi także być taka wielkość
populacji
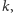 dla której następuje całkowite zahamowanie wzrostu, czyli
dla której następuje całkowite zahamowanie wzrostu, czyli
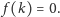 Najprostszą funkcją spełniającą wymienione założenia jest
liniowa funkcja
Najprostszą funkcją spełniającą wymienione założenia jest
liniowa funkcja
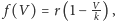 gdzie
gdzie
 oznacza teraz
maksymalny współczynnik wzrostu populacji.
oznacza teraz
maksymalny współczynnik wzrostu populacji.
Otrzymamy w ten sposób następujący rozbudowany model drapieżnik–ofiara:
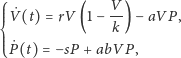 | (2) |
gdzie zmienne i parametry (oprócz
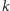 ) mają taką samą interpretację jak
dla (1), zaś parametr
) mają taką samą interpretację jak
dla (1), zaś parametr
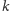 oznacza pojemność środowiska dla
gatunku ofiar.
oznacza pojemność środowiska dla
gatunku ofiar.
Zajmiemy się teraz analizą jakościową przebiegu rozwiązań. W tym celu
narysujemy portret fazowy modelu, z którego odczytamy globalne zachowanie
rozwiązań. Zaczniemy od wyznaczenia izoklin zerowych, czyli krzywych,
na których zerują się prawe strony pierwszego (dla zmiennej
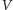 )
i drugiego (dla zmiennej
)
i drugiego (dla zmiennej
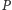 ) równania. Izokliny mają postać:
) równania. Izokliny mają postać:
- dla zmiennej
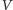 :
:
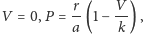
- dla zmiennej
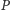 :
:
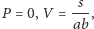
co przedstawiają rysunki na marginesie.
Stany stacjonarne odczytujemy jako punkty przecięć odpowiednich izoklin.
Zauważmy, że gdy
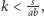 to jeden z punktów przecięcia pojawia się
poza pierwszą ćwiartką układu (w ćwiartce czwartej, na przedłużeniu
odpowiednich prostych), a zatem wykracza poza dziedzinę rozważań
modelu, zaś gdy
to jeden z punktów przecięcia pojawia się
poza pierwszą ćwiartką układu (w ćwiartce czwartej, na przedłużeniu
odpowiednich prostych), a zatem wykracza poza dziedzinę rozważań
modelu, zaś gdy
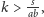 to punkt ten mieści się w pierwszej
ćwiartce. Ponieważ żadna z tych nierówności nie jest wyróżniona
przez nasz model, analiza musi obejmować dwa przypadki. Przypadek
trzeci,
to punkt ten mieści się w pierwszej
ćwiartce. Ponieważ żadna z tych nierówności nie jest wyróżniona
przez nasz model, analiza musi obejmować dwa przypadki. Przypadek
trzeci,
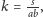 jest przypadkiem granicznym, w którym punkty
jest przypadkiem granicznym, w którym punkty
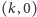 i
i
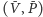 sklejają się – jest on tożsamy z sytuacją,
gdy
sklejają się – jest on tożsamy z sytuacją,
gdy
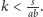
Możemy wykazać, że są dwa lub trzy stany stacjonarne o następującym charakterze:
-
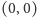 jest siodłem,
jest siodłem,
-
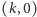 jest węzłem stabilnym asymptotycznie lub siodłem,
jest węzłem stabilnym asymptotycznie lub siodłem,
-
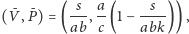 istnieje o ile
istnieje o ile
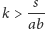 i jest węzłem lub ogniskiem stabilnym.
i jest węzłem lub ogniskiem stabilnym.
Na podstawie tych rysunków możemy naszkicować portrety fazowe, które
przedstawiamy na sąsiedniej stronie (w przypadku
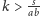 punkt
punkt
 narysowany został jako ognisko, choć może być też
węzłem).
narysowany został jako ognisko, choć może być też
węzłem).
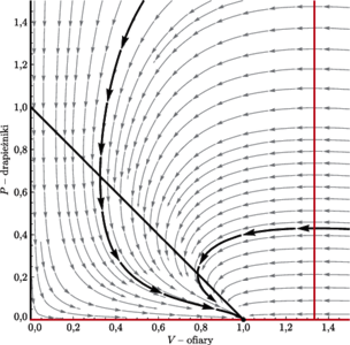
Portret fazowy w przypadku, gdy
Portret fazowy w przypadku, gdy
|
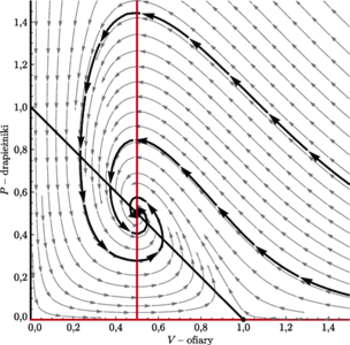
i w przypadku, gdy
i w przypadku, gdy
|
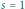 ; różnią się wyłącznie wartością
współczynnika
; różnią się wyłącznie wartością
współczynnika
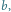 przy czym na wykresie lewym
przy czym na wykresie lewym
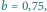 zaś na prawym
zaś na prawym
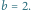 Na obu portretach wyróżnione zostały po dwie przykładowe orbity:
jedna przechodząca przez punkt
Na obu portretach wyróżnione zostały po dwie przykładowe orbity:
jedna przechodząca przez punkt
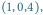 druga przez punkt
druga przez punkt
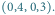
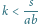
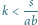
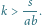
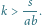
Patrząc na te portrety fazowe, widzimy ciekawe rzeczy.
- Osie układu są orbitami.
- Jeśli punkt
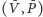 jest poza dziedziną lub skleja się
z
jest poza dziedziną lub skleja się
z
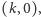 to izokliny
dzielą przestrzeń fazową na trzy obszary – nazwijmy je
to izokliny
dzielą przestrzeń fazową na trzy obszary – nazwijmy je
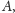
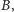
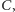 licząc od lewej. Rozwiązanie zaczynające
się w
licząc od lewej. Rozwiązanie zaczynające
się w
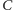 musi przejść do
musi przejść do
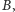 bo w przeciwnym
razie współrzędna
bo w przeciwnym
razie współrzędna
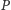 uciekałaby do nieskończoności,
zaś w obszarach
uciekałaby do nieskończoności,
zaś w obszarach
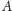 i
i
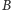 obie współrzędne
są monotoniczne i ograniczone, a więc zbieżne. Zatem punkt
stacjonarny
obie współrzędne
są monotoniczne i ograniczone, a więc zbieżne. Zatem punkt
stacjonarny
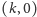 jest rozwiązaniem globalnie stabilnym.
jest rozwiązaniem globalnie stabilnym.
- Gdy punkt
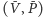 jest wewnątrz dziedziny
jest wewnątrz dziedziny
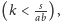 to jest on lokalnie stabilny. Stosując odpowiednie
metody (np. metodę funkcji Lapunowa), możemy też wykazać,
że jest on globalnie stabilny.
to jest on lokalnie stabilny. Stosując odpowiednie
metody (np. metodę funkcji Lapunowa), możemy też wykazać,
że jest on globalnie stabilny.
Wnioski
Po przeanalizowaniu układu (2) otrzymaliśmy jeden punkt globalnie stabilny:
- punkt
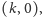 o ile
o ile
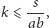
- punkt
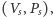 o ile
o ile
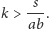
Każdy z tych punktów jest zbiorem granicznym, do którego zbiegają wszystkie inne orbity układu, poza orbitami biegnącymi wzdłuż układu współrzędnych. Znaczy to, że dowolne rozwiązanie z warunkiem początkowym o obu współrzędnych dodatnich, czyli zaczynające się od dodatnich populacji tak drapieżnika, jak i ofiary, będzie z upływem czasu zbiegało do rozwiązania stacjonarnego, jednego z dwóch powyższych – do którego, to zależy od wartości parametrów układu.
Pamiętamy, że chcieliśmy porównać dynamikę populacji drapieżnika
stałocieplnego ze zmiennocieplnym w tym samym środowisku. Skoro tak,
to możemy przyjąć, że współczynniki
 oraz
oraz
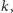 jako opisujące
populację ofiar, są zadane z góry przez model (albo np. pomiary terenowe).
Natomiast dla drapieżników – skoro chcemy porównywać dwa różne
gatunki, to będziemy porównywali model z dwoma zestawami parametrów:
jako opisujące
populację ofiar, są zadane z góry przez model (albo np. pomiary terenowe).
Natomiast dla drapieżników – skoro chcemy porównywać dwa różne
gatunki, to będziemy porównywali model z dwoma zestawami parametrów:
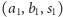 dla pierwszego drapieżnika oraz
dla pierwszego drapieżnika oraz
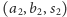 dla
drugiego drapieżnika.
dla
drugiego drapieżnika.
Zauważmy, że wystarczy manipulacja współczynnikiem
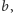 aby wybrać,
czy
aby wybrać,
czy
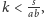 czy
czy
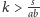 – im większe
– im większe
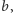 tym mniejszy
ułamek po prawej stronie. Parametr
tym mniejszy
ułamek po prawej stronie. Parametr
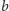 w modelu (2) opisuje część
energii pozyskanej z upolowanej ofiary przeznaczoną na rozród drapieżnika.
Jak już było zaznaczone we wstępie, zwierzęta zmiennocieplne mają
kilkakrotnie mniejsze zapotrzebowanie na energię niż stałocieplne,
gdyż nie muszą utrzymywać stałej temperatury ciała i szybkiego tempa
metabolizmu, co oznacza, że relatywnie więcej energii mogą przeznaczyć
na reprodukcję. W kontekście omawianego modelu (2) oznacza to większą
wartość współczynnika
w modelu (2) opisuje część
energii pozyskanej z upolowanej ofiary przeznaczoną na rozród drapieżnika.
Jak już było zaznaczone we wstępie, zwierzęta zmiennocieplne mają
kilkakrotnie mniejsze zapotrzebowanie na energię niż stałocieplne,
gdyż nie muszą utrzymywać stałej temperatury ciała i szybkiego tempa
metabolizmu, co oznacza, że relatywnie więcej energii mogą przeznaczyć
na reprodukcję. W kontekście omawianego modelu (2) oznacza to większą
wartość współczynnika
Można tę sytuację interpretować w ten sposób, że dwa różne przypadki portretów fazowych, a więc i różne możliwe zachowania rozwiązań, odpowiadają dwóm różnym gatunkom drapieżników, przy czym przypadek pierwszy, gdy rozwiązania układają się jak na rysunku z lewej, oznacza, że mamy do czynienia z drapieżnikiem stałocieplnym, zaś gdy wyglądają jak na rysunku z prawej, z drapieżnikiem zmiennocieplnym. To uzasadnia, zgodnie z przypuszczeniami Flannery’ego, nieuchronność zagłady australijskich drapieżnych torbaczy i jednoczesne przetrwanie stabilnej populacji mięsożernych gadów.
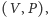 gdzie rozwiązania są
krzywymi opisanymi lokalnie jako funkcje
gdzie rozwiązania są
krzywymi opisanymi lokalnie jako funkcje
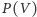 lub
lub
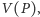 a przebieg w czasie zaznaczamy strzałkami, których kierunek wyznaczony jest
przez wektor
a przebieg w czasie zaznaczamy strzałkami, których kierunek wyznaczony jest
przez wektor