Kącik biologiczny
Skąd się biorą geny?
Około 40 milionów lat temu u wspólnego przodka małp Starego Świata (do których należą naczelne) miało miejsce niezwykłe wydarzenie. Umożliwiło ono nam zupełnie nowe spojrzenie na świat – dosłownie.
Widzenie barw oparte jest na światłoczułych białkach – opsynach, które, zależnie od składu aminokwasowego, są wrażliwe na różne długości fali. Większość małp Nowego Świata (np. kapucynki) ma dwa geny opsyn: jeden z nich, umiejscowiony na autosomie, koduje białko wrażliwe na fale krótkie (SW), a drugi, kodujący białko wrażliwe na fale średnie i długie (MW/LW, zależnie od sekwencji), znajduje się na chromosomie X. Stąd samce tych małp widzą wyłącznie dwubarwnie, natomiast samice – albo dwu- (jeśli na obu chromosomach X mają taką samą wersję opsyny MW/LW), albo trójbarwnie (jeśli wersje te się różnią)!
Mniej więcej 40 milionów lat temu, po oddzieleniu się małp Nowego Świata od linii prowadzącej do człowieka, gen opsyny MW/LW uległ duplikacji. Oba nowe geny szybko zróżnicowały się tak, że każdy z nich kodował białko wrażliwe na inną długość fali. Odtąd wszystkie małpy mają trzy geny opsyn – SW, MW i LW: była to bardzo dobra wiadomość dla samców, które wreszcie mogły widzieć świat trójkolorowo.
Ten niezwykły przypadek ilustruje najczęstszy sposób powstawania genów. Kiedy w genomie pojawia się kopia istniejącego już genu, presja selekcyjna, utrzymująca funkcję białka, działa tylko na jedną z nich. Druga dzięki temu bezkarnie mutuje, co czasem prowadzi do uzyskania nowej funkcji. Ewolucja nie stwarza z niczego, może tylko modyfikować istniejące rozwiązania.
W ten sposób mogą powstawać tzw. rodziny genów – grupy podobnych sekwencji powstałe na drodze duplikacji i różnicowania. Przykładem takiej rodziny są globiny, które wszystkie wyewoluowały w ciągu 800 milionów lat na drodze kolejnych duplikacji, fuzji i modyfikacji funkcji. Mioglobina, mięśniowe białko wiążące tlen, i 13 globin krwi u człowieka są takimi właśnie kuzynami.
Czasem duplikacje nie dotyczą całych genów, a jedynie ich fragmentów. Mogą być one tasowane niczym talia kart w obrębie jednego genu, albo przenoszone do innego. Znane są geny – układanki z kilku różnych sekwencji. Gen kodujący jedno z białek regulujących krzepnięcie krwi, tkankowy aktywator plazminogenu, zbudowany jest z czterech części. Pierwsza jest kawałkiem fibronektyny (wiążącej włókna białek tworzących skrzep), druga pochodzi z nabłonkowego czynnika wzrostu, a dwie ostatnie – z plazminogenu (formy enzymu rozpuszczającego skrzep). W wyniku takiego tasowania zupełnie różne i niespokrewnione geny mogą mieć bardzo podobną sekwencję.
W analizie pokrewieństwa między organizmami musimy rozróżniać sekwencje, które pojawiły się w wyniku duplikacji w obrębie genomu jednego gatunku (czyli tzw. paralogi) oraz te, które są wynikiem rozdzielenia gatunków (ortologi).
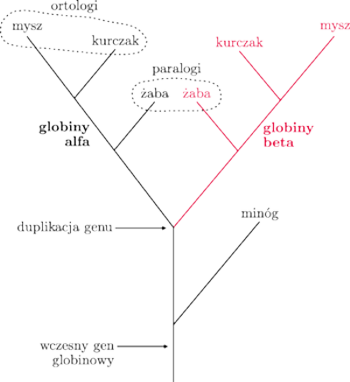
Z rysunku widać, że jeden gen może mieć więcej niż jednego ortologa u drugiego gatunku (zarówno globiny alfa, jak i beta, żaby są ortologami w stosunku do globinowego genu minoga), a także, że „bycie ortologiem” nie jest przechodnie (globiny alfa i beta myszy są ortologami globiny minoga, ale nie dla siebie).
Wyobraźmy sobie, że chcemy oszacować wiek ostatniego wspólnego przodka (OWP) człowieka i wielbłąda na podstawie liczby mutacji nagromadzonych w ich genach. W tym celu musimy porównać geny ortologiczne, np. gen A człowieka i odpowiadający mu gen wielbłąda. Jak znaleźć ortolog? Wśród genów wielbłąda jest wiele podobnych do genu A, najbardziej podobny jest gen B. Czy A i B to ortologi? Tego nie wiemy – zarówno orto-, jak i paralogi mogą być bardzo podobne. Genom wielbłąda nie został jeszcze zsekwencjonowany i wśród dostępnych sekwencji może w ogóle nie być ortologa. Może być też tak, że wielbłąd utracił ortolog genu A.
W praktyce przeszukujemy jeszcze raz genom, tym razem genom człowieka sekwencją B. Jeżeli najbardziej podobną do niej sekwencją jest A, to uznajemy geny za ortologi i na ich podstawie możemy wnioskować o OWP wielbłąda i człowieka. Nie daje nam to pewności, ale status sekwencji A i B możemy potwierdzić, przeszukując w podobny sposób kolejne genomy. W każdym przypadku dwukierunkowe przeszukania powinny prowadzić do tych samych sekwencji. A jeśli te geny to nie ortologi i porównujemy np. podobne do siebie hemoglobinę człowieka i mioglobinę wielbłąda?
Wtedy okazałoby się, że OWP tych gatunków żył około 800 milionów lat temu – bo wtedy właśnie nastąpiło rozdzielenie genów mioglobiny i hemoglobiny. A 800 milionów lat temu wielokomórkowe życie dopiero nabierało rozpędu… Naukowcom wciąż pozostaje jeszcze znalezienie odpowiedzi na pytanie, skąd wziął się pierwszy gen. Ale to jest już zupełnie inna historia...